chromosome
Cet article fait partie du DOSSIER consacré à la
Structure en forme de bâtonnet du noyau cellulaire, constituant le support physique de l'hérédité.
Les chromosomes, qui apparaissent dans le noyau de la cellule au moment de la division (mitose ou méiose),
résultent de la condensation de la chromatine (réseau diffus d'ADN et
de protéines). La structure de ces bâtonnets, qui constituent le support
des caractères héréditaires, est aujourd'hui l'objet de recherches
considérables à l'échelle mondiale.
La découverte des chromosomesEntre
les années 1870 et 1880, les cytologistes donnent le nom de chromatine
au réseau filamenteux colorable qu'ils observent dans le noyau, puis
celui de mitose au mécanisme de division cellulaire, car il est caractérisé par l'apparition de filaments (en grec,
mitos).
En 1888, ces filaments reçoivent le nom de chromosomes, en raison de
leur aptitude à se colorer fortement en présence de colorants basiques
(du grec
khrôma, couleur, et
sôma, corps).
Chromatine et chromosomes ne sont que deux aspects différents d'une même entité.Au début du XX
e s.,
le parallélisme est fait entre le comportement des chromosomes au cours
des divisions menant à la formation des gamètes (méiose) et les règles
de transmission des caractères héréditaires, définies par Gregor Mendel
quarante ans plus tôt (les travaux de ce moine scientifique avaient été
publiés en 1866, puis oubliés).
De cette constatation naît
la théorie chromosomique de l'hérédité (les chromosomes sont les supports physiques de l'information héréditaire). C'est l'Américain Thomas Hunt Morgan
qui, en 1920, consacre l'union entre génétique et cytologie : ses
travaux sur les chromosomes et le patrimoine génétique de la drosophile
(mouche du vinaigre) ont permis l'établissement des représentations de
l'ensemble des chromosomes d'une cellule, ou
caryotypes, sur lesquelles les chromosomes sont rangés par paires. Les gènes,
terme introduit par Wilhelm Johannsen en 1909 pour qualifier les
éléments transmetteurs de l'hérédité, occupent une position fixe sur les
chromosomes. Depuis, la composition chimique des chromosomes et la
structure en double hélice de l'ADN
ont été définies, mais ce n'est qu'à partir de 1974 que l'agencement
des différentes molécules les unes par rapport aux autres a été établi.
La morphologie des chromosomesL'examen des cellules animales et végétales en division montre que
chaque chromosome a une taille et une forme propres ; à de rares exceptions près,
leur nombre est pair.
L'observation est effectuée sur des cellules dont la division a été
bloquée, à l'aide de colchicine (alcaloïde extrait du colchique), au
stade de la métaphase (deuxième des quatre stades que comporte le
processus de division cellulaire). À ce moment, les chromosomes sont
séparés dans le sens longitudinal en deux chromatides, ou chromosomes
fils, réunis par un centromère (constriction primaire).
Dans toutes les cellules somatiques, ou non reproductrices,
chaque type de chromosome existe en deux exemplaires : ils sont dits
homologues. En définissant par
n le nombre de chromosomes différents, les généticiens ont qualifié de
diploïdes ces cellules qui possèdent 2
n chromosomes. En revanche, les cellules reproductrices, ou gamètes, qui ne contiennent que
n chromosomes sont
haploïdes.
Le nombre de chromosomesAu moment de la fécondation, la fusion des deux gamètes (spermatozoïde et ovule) rétablit le nombre de chromosomes (
n +
n = 2
n) dans la cellule œuf ou zygote. Le nombre de paires de chromosomes dans les cellules humaines, 23 au total (2
n = 46), a été établi en 1956 par Joe Hin Tijo et Albert Levan. Il est important de remarquer que
le nombre chromosomique d'une espèce n'a aucun rapport avec son degré d'évolution, ni même avec sa taille, puisque l'amibe possède plusieurs centaines de chromosomes.
L'utilisation
expérimentale de substances comme la colchicine permet de provoquer des
anomalies numéraires en déréglant le processus de la mitose. Les
espèces créées possèdent un caryotype dont les chromosomes sont en
quadruple exemplaire (cellules tétraploïdes, à 4
n), voire plus.
De nombreuses plantes cultivées (blé, rose, tabac, dahlia, etc.), qui
ont fait l'objet de telles manipulations génétiques, sont caractérisées
par des fleurs, des graines et une taille générale supérieures à la
normale.
Les chromosomes sexuels
Chromosomes sexuels
Chez
l'espèce humaine, ce sont ceux de la vingt-troisième paire sur un
caryotype. Chez la femme, cette paire est composée de deux chromosomes
identiques, nommés X. Chez l'homme, elle est composée de deux
chromosomes de taille différente : l'un, ressemblant à ceux de la femme,
est un chromosome X ; l'autre, plus court, est le chromosome Y. Appelés
aussi
hétérochromosomes, les chromosomes sexuels
déterminent le sexe de l'individu, par opposition aux 22 autres paires, les
autosomes,
qui définissent les caractères physiques et psychologiques de
l'individu. Chez d'autres espèces, telle la mouche, la femelle est XY,
alors que le mâle est XX.
D'autre
part, les cellules d'une femme sont caractérisées, entre deux
divisions, par la présence dans le noyau d'un grain de chromatine,
correspondant à l'un de ses chromosomes sexuels. Ce grain, aussi appelé
chromatine sexuelle ou corpuscule de Barr, sert à diagnostiquer le sexe
vrai ou des anomalies sexuelles.
Des chromosomes particuliersLe « chromosome » bactérienGlobalement,
le matériel génétique d'une bactérie est comparable à celui des
eucaryotes. Il est également constitué d'ADN et de protéines qui, sans
être rigoureusement identiques, conservent les mêmes fonctions. Le
génome principal d'une bactérie est représenté par
une molécule d'ADN de forme circulaire qui, par analogie avec la structure du génome des eucaryotes, est
souvent appelée « chromosome bactérien », bien qu'il ne s'agisse pas
d'un chromosome à proprement parler.
Nombre
de bactéries ne possèdent qu'un seul « chromosome », mais il n'est pas
rare d'en observer plusieurs, surtout avant la période de division. La
souche de laboratoire d'
Escherichia coli, un bacille
particulièrement étudié dans les domaines de la génétique et de la
biologie moléculaire, possède un chromosome de 1,3 mm de circonférence,
contenant environ 4,7 millions de paires de bases. Son génome a été
entièrement séquencé (1997). En 1997, une équipe internationale de
plusieurs dizaines de chercheurs a publié la séquence complète des
4 214 810 paires de bases constituant le génome de la bactérie
Bacillus subtilis, et déterminé ses 4 100 gènes codant pour des protéines. Depuis, de nombreuses autres bactéries (
Helicobacter pylori, Haemophilus influenzae, Saccharomyces cerevisiae, Mycoplasma pneumoniae, etc.) et archaebactéries (
Archaeoglobus fulgidus, Methanococcus thermoautotrophicum, Aquifex aeolicus, etc.) ont déjà vu ainsi leur génome entièrement séquencé (il est
d'ailleurs remarquable que ces données soient librement accessibles sur
Internet).
Les bactéries possèdent également, en nombre variable,
d'autres fragments de génome de taille beaucoup plus réduite, représentant un centième du chromosome principal. Ces fragments, circulaires eux aussi, sont appelés
plasmides et
se répliquent de façon indépendante. L'un d'entre eux, le facteur F, confère le caractère mâle à la bactérie porteuse. Il permet la
conjugaison et le transfert de matériel génétique d'une bactérie à l'autre, tout
particulièrement lorsqu'il s'intègre au chromosome principal. En
général, les plasmides ne portent pas d'information génétique
essentielle au métabolisme de la bactérie. C'est toutefois dans ces fragments d'ADN que sont contenus les gènes conférant la résistance aux antibiotiques.
Les chromosomes géantsChez
certaines espèces, on peut observer des chromosomes de très grande
taille, environ près de cent fois celle d'un chromosome de dimension
moyenne. Ces chromosomes géants, ou polyténiques, ont été trouvés chez
des protozoaires ciliés et dans les cellules du tube digestif ou de
glandes annexes de diptères (mouches), comme la drosophile. Ils sont
constitués par l'association des deux chromosomes homologues, eux-mêmes
formés d'un millier de nucléofilaments, accolés les uns aux autres après
la réplication. Une alternance de bandes sombres et claires est
particulièrement visible sur cette structure ; elle correspond aux
différences de condensation du nucléofilament. Toutes ces spécificités
ont fait des chromosomes géants un objet privilégié pour les études
génétiques, et notamment pour la cartographie des gènes chez la
drosophile.
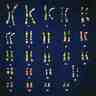
Le caryotype
Caryotype
Le
caryotype est l'ensemble des chromosomes d'une cellule ou d'un
individu, et spécifique de l'espèce à laquelle il appartient (le nombre
de chromosomes d'une espèce est constant).
Établissement du caryotypeChez
l'homme, le caryotype est réalisé à partir de cellules sanguines, les
globules blancs, qui se divisent assez facilement en culture. Après un
prélèvement de sang par ponction veineuse, les cellules sont déposées
sur un milieu de culture contenant des éléments nutritifs, des
antibiotiques (pour éviter leur contamination et leur destruction par
des bactéries) et des substances activant la mitose.
Trois jours plus tard,
les mitoses sont bloquées à l'aide de colchicine,
qui empêche la mise en place du fuseau. Les cellules, soumises à un
choc hypotonique qui les fait éclater, sont ensuite fixées par des
mélanges à base d'alcool, de chloroforme et d'acide acétique. Enfin,
les chromosomes sont colorés, photographiés, identifiés grâce aux clichés, puis classés.
La formule chromosomiqueLe
chromosome Y est certainement le chromosome humain le plus variable du
patrimoine génétique de l'homme. Sa longueur varie selon les
populations : par exemple, il est souvent plus grand chez les Asiatiques
que chez les Européens. Certains chromosomes possèdent des
constrictions secondaires, sortes d'allongements supplémentaires ; le
chromosome 9, à l'état normal, en possède une sur son bras long, mais,
chez certains individus, elle se situe sur le bras court de ce
chromosome. La constriction des chromosomes 1 et 16 peut être de
longueur variable.
De
telles variations sont également visibles chez les animaux, comme les
rongeurs, qui montrent de nombreux exemples de variations
intraspécifiques de la formule chromosomique. Certains individus
possèdent même un nombre de chromosomes inférieur ou supérieur à la
normale. Des différences existent chez des espèces extrêmement proches
les unes des autres, tant sur le plan de la physiologie que sur celui de
l'anatomie, et pour lesquelles aucune différence chromosomique majeure
ne pourrait être attendue. La stabilité du caryotype n'est donc que
relative, et cela quelle que soit la position de l'être vivant dans la
classification zoologique. Toutefois ces altérations sont peu fréquentes
dans une population donnée.
Pour
de nombreux auteurs, ces variations de la formule chromosomique peuvent
être dans certains cas, au même titre que les mutations ponctuelles des
gènes, un élément d'évolution d'une espèce – tout au moins lorsque ce
changement est susceptible d'induire des changements de phénotype et
lorsque ceux-ci restent compatibles avec la survie de l'individu.
Les aberrations chromosomiques
Aberration chromosomique
Il
arrive que les altérations du stock chromosomique soient profondes et
entraînent de graves problèmes de santé pour l'individu qui les porte.
Ces anomalies chromosomiques sont
soit de type numérique, soit de type structural, et peuvent affecter tous les chromosomes.
Les
anomalies de type numérique sont principalement dues à une mauvaise
séparation des chromosomes appariés au cours de la mitose ou de la
méiose. Lorsque le caryotype comprend un multiple exact du nombre
haploïde (3
n ou 4
n, par exemple), la cellule ou l'organisme sont dits
polyploïdes. Si le nombre n'est pas un multiple exact, on parle d'
aneuploïdie.
La trisomie 21 (ou syndrome de Down) est un exemple d'aberration de ce
type : elle est caractérisée par la présence d'un chromosome
surnuméraire dans la paire 21. La principale cause de l'aneuploïdie est
une distribution inégale du stock chromosomique lors de la formation des
cellules sexuelles.
Les
aberrations de la structure chromosomique résultent d'une cassure d'un
chromosome suivie d'une reconstruction anormale. Elles peuvent
apparaître à la suite de divers mécanismes. La délétion (perte pure et
simple d'un fragment du chromosome) peut concerner sa partie terminale
ou une portion comprise entre deux points de rupture. La duplication
aboutit à l'intégration d'un fragment supplémentaire de chromosome tout
en créant une délétion dans un autre chromosome. Dans d'autres cas, si
la cassure d'un chromosome est mal réparée, le fragment peut être
« ressoudé » à l'envers (inversion) ou intégré en un tout autre site
(translocation).
Voir aussi : hérédité, aberration chromosomique, délétion, gène, noyau.
La structure des chromosomesLes chromosomes et la chromatine sont constitués de protéines et d'acides nucléiques (ADN). L'ARN
n'intervient pas dans la structure même, car l'information portée par
la molécule d'ADN est transcrite sous forme d'ARN qui, après être passé
du noyau vers le cytoplasme, est traduit en protéine.
L'ADN chromosomique
A.D.N.
La
quantité d'ADN contenue dans une cellule est constante pour une espèce
vivante donnée, exception faite des cellules reproductrices dont le
stock est divisé par 2. Cette quantité a été estimée à 0,4 pg chez la
mouche et à 5,86 pg chez le chien, par exemple. Pour l'homme, les
estimations ont donné 7,3 pg pour une longueur totale de
2,36 m.
Si l'on veut avoir une idée plus réelle des proportions de la molécule
d'ADN, il suffit de multiplier ses dimensions un million de fois. Dans
ce cas, les 46 brins d'ADN complètement déroulés des chromosomes humains
mis bout à bout formeraient un fil de 2 mm de diamètre pour une
longueur de plus de 2 000 km !
La
molécule d'ADN est formée d'un double enchaînement ordonné de
nucléotides complémentaires (fibre bicaténaire, ou duplex) de 2 nm de
diamètre. Cet arrangement définit sa structure primaire globalement
linéaire; mais par les propriétés physicochimiques de ses composants,
l'ADN adopte une configuration en double hélice qui correspond à sa
structure secondaire. Selon les cas, on compte entre 9 et 11 bases par
tour d'hélice. Cette molécule bicaténaire est relativement rigide le
long de son axe et subit, par conséquent, de nombreuses contraintes dans
l'architecture des chromosomes. Chez l'homme, un chromosome peut
mesurer une dizaine de micromètres de long pour 0,5 mm de diamètre :
l'ADN est donc fortement compacté au sein d'une structure qui inclut
aussi de nombreuses protéines.
Les protéines chromosomiques
Chromosome
Elles appartiennent à deux groupes, celui des
histones, très homogène, et celui des
non-histones, qui comportent plusieurs centaines de types différents.
Par
leur richesse en acides aminés, telles la lysine et l'arginine, qui à
eux seuls peuvent constituer près de 25 % de la chaîne polypeptidique,
les histones sont des protéines basiques. Elles ont une forte affinité
entre elles, mais aussi avec les acides nucléiques comme l'ADN
et avec les protéines non histones. Si l'on considère leur structure
primaire, ou séquence d'acides aminés, on constate une remarquable
constance, même pour des espèces d'eucaryotes fort éloignées. Ce qui
laisse à penser que le rôle de ces protéines est identique pour toutes
les espèces et que la moindre anomalie de séquence doit être mortelle
pour l'organisme. Les protéines histones sont réparties en cinq types
majeurs : H1, H2A, H2B, H3 et H4.
Les
protéines non histones forment un groupe très hétérogène du fait de la
diversité des rôles qu'elles assurent. Certaines d'entre elles
interviennent, tout comme les histones, dans l'architecture du
chromosome. La myosine et l'actine, par exemple, qui sont également les
protéines contractiles fondamentales du muscle, pourraient entrer en
action lors de la condensation et la décondensation des chromosomes. On
trouve par ailleurs des protéines, dont le rôle est d'assurer la
réplication et la réparation des brins d'ADN, et toutes les enzymes,
substances protéiniques, nécessaires à la transcription des gènes en
ARN.
La structure de base des chromosomes
Crossing-over
Quel que soit l'aspect qu'il présente, la structure de base du chromosome est la
fibre nucléosomique.
Révélée à la fin des années 1950 par des diagrammes de diffraction des
rayons X, cette structure ressemble à un chapelet de perles dont le fil
connecteur serait l'ADN et les perles des groupements d'histones. C'est
Arthur Kornberg qui en 1974, sur la base de clichés réalisés en
microscopie électronique, proposa un modèle pour décrire cet
arrangement. Le nucléosome, unité de base, serait constitué par un
assemblage protéique contenant 8 histones, identiques deux à deux
(2 × H2A, 2 × H2B, 2 × H3, 2 × H4). Le filament d'ADN s'enroulerait
autour de cette structure en faisant deux tours de spire à la manière
d'un fil s'enroulant sur un court cylindre (110 Å de diamètre sur 57 Å
de hauteur) ; la continuité du filament d'ADN assure également le lien
entre les différents nucléosomes.
L'organisation
du nucléofilament dans la chromatine est encore plus complexe. Des
filaments environ trois fois plus épais que la fibre nucléosomique (soit
d'un diamètre de près de 30 nm) seraient le résultat de la condensation
de la fibre nucléosomique. Deux modèles ont été proposés pour en
décrire l'organisation : en solénoïde et en superboules. Dans le
premier, la fibre nucléosomique s'enroulerait en une spirale dont chaque
tour serait formé de 6 nucléosomes. Dans le second, la fibre
chromosomique serait formée par la juxtaposition de boules contenant
12 nucléosomes. La stabilité de ces deux structures serait assurée par
les protéines histones de type H1.
Un
des stades privilégiés de la division cellulaire pour étudier les
chromosomes est la métaphase. En effet, à ce moment, ils sont
complètement individualisés en chromatides, et la condensation, amorcée
au début de la phase de division, est maintenant terminée. Les
chromatides formées correspondent à un degré d'organisation
supplémentaire de la fibre chromosomique décrite précédemment.
Dans
cette organisation, les protéines non histones jouent un rôle très
important. On a pu isoler et purifier une trentaine d'entre elles, ce
qui ne représente qu'un millième de la quantité d'histones constituant
le chromosome. Ces protéines forment le squelette longiligne qui donne
sa forme générale à la chromatide ; la fibre chromosomique, qui
s'enroulerait autour de cet axe, formerait au préalable des sortes de
petites pelotes, les microconvules, de 52 nm de diamètre, contenant
l'ADN. Ce nouveau chapelet se disposerait alors autour de l'axe de
protéines non histones. Une estimation avance que le chromosome humain
le plus grand pourrait être composé de plus de 4 000 de ces
microconvules.
La duplication des chromosomesToute division cellulaire (mitose, première division de la méiose) est précédée d'une
duplication de l'information génétique, afin que celle-ci puisse être transmise dans son intégralité aux cellules filles. Cette duplication est assurée par la
réplication de l'ADN,
qui aboutit à la formation de deux longues molécules linéaires en tous
points semblables. Elle s'effectue selon un modèle semi-conservatif,
dans lequel chaque brin de la double hélice engendre un brin
complémentaire puis s'y associe. La réplication est un phénomène
biochimique très complexe nécessitant la participation de nombreuses
enzymes dont le rôle est de dérouler le filament d'ADN, de séparer les
deux brins, de synthétiser les brins complémentaires et, enfin, de
reconstituer la structure native des brins fils. (→ ADN)
La parenté chromosomique de l'hommeLes
scientifiques ont généralement recours à la morphologie, à l'anatomie
et à la physiologie pour déterminer les liens évolutifs entre
différentes espèces. Mais ces liens peuvent être établis par l'analyse
de la formule chromosomique. Depuis Darwin, les biologistes ont admis
que l'homme et les grands singes (chimpanzé, gorille, orang-outan…) ont
une parenté commune ; elle a été confirmée par l'étude de leur formule
chromosomique.
L'homme
a 23 paires de chromosomes, contre 24 chez les grands singes. Une
analyse plus fine a montré que 13 des paires humaines sont
rigoureusement identiques à celles du chimpanzé et que les paires
restantes ne se distinguent que par de subtils remaniements. Quant au
chromosome 2 de l'homme, il correspond tout simplement à la fusion de
deux chromosomes du chimpanzé.
Les
scientifiques se demandent à présent pourquoi des différences aussi
minimes peuvent se traduire par des divergences morphologiques et
comportementales aussi importantes.



 جنسيتي
جنسيتي

 الجنس
الجنس عدد المساهمات
عدد المساهمات نقاط
نقاط العمر
العمر